CARTAUCSD@CARTAUCSD Why don’t we live longer? Because evolution is more concerned with helping us produce healthy children.
1:02 To answer the question why we die...the principle of natural selection that drives evolution and it has to do with the cost-benefit analysis of how many limited resources you put into organisms. So, obviously natural selection is going to select for infants and toddlers and babies to be well cared for...have super regenerative powers to keep their bodies going in order to get the genes into the next generation (get them up to reproductive age) and so on. So, we see cells that divide very rapidly in infants and babies in a little cut...whereas someone my age, when I get cut, takes much longer to heal. The question is why wouldn't evolution just make it so that I (now in my early sixties) can't just keep going to 200, 300 and the answer is there's no reason for it because after I've brought my own offspring into reproductive age and they've brought their offspring into reproductive age I'm really of no use anymore. I can serve a useful purpose as a parent, of course, bringing my genes up and then a useful purpose as a grandparent to help my offspring bring their offspring up to reproductive age, but beyond that...there's no sense in pouring anymore resources into great-great-great-great-great grandparents because the genes in the little infant are already going to be well taken care of...this is just how natural selection operates. So, in short, we die so that our future generations may live because there are limited resources.
1:08 We know that the human body is so complex that if you live to 150 or 200 there may be other things that happen. We don't even know yet. I mean, most people 500 years ago had no idea about Alzheimer's other than the handful of people that seemed to have lost their memories, but now we realize because so many of us live that long.
The occurrence of gene-culture coevolution is fundamental to the unity of science and humanities. Consider, as an example, the process of aging. Why must we die? More generally, why does each species in turn, ours included and each hereditary strain within each species, of which humanity has plenty, possess a characteristic lifespan? If you seek a long-lived dog as a companion, as opposed say to a sheep herder or wild-boar hunter, you would be better off with a Chihuahua (lifespan twenty years) than with a Great Dane (lifespan six years). Plants also have programmed lifespans. Some northern conifers live on average about a century, magnolias a century-and-a-half, and sequoias and pines of the Southwestern United States for as long as several millennia. But they, too, age and die eventually. What could be more important to both science and the humanities than the human life cycle and preordained human life-spans?
The prevailing theory in evolutionary biology to explain programmed aging and death recognizes that each species of plant and animal has evolved a lifestyle in which most individuals die from external causes - disease, accident, congenital defect, malnutrition, murder, war - well short of their maximum potential longevity. Such was severely the rule in Paleolithic times, when few people reached an age of fifty years. As a result of this idiosyncratic pre-longevity death rate, which is improved but applies to most humans today, natural selection has front-loaded vigor and reproductive drive.It has programmed the vital physiology and mental state of youth in the youngest adults, while disfavoring older adults. It has bet, so to speak, against investment in middle and old age.
With the dawn of Neolithic civilization, and advent of agriculture and food storage, along with an easing of external mortality, conditions have changed in a way that redirects natural selection along the human life cycle. Thanks to the weakening of Paleolithic mortality factors through cultural evolution, average longevity is increasing and the age of reproduction expanding up to the age of menopause.
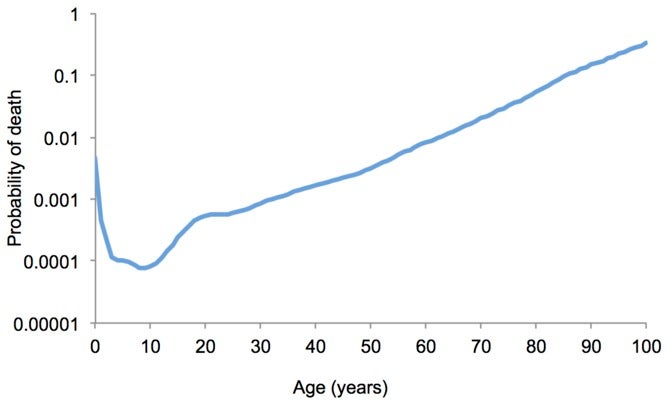
In 1952, the British biologist Peter Medawar formalized the first evolutionary theory of aging, terming it the mutation-accumulation model. This theory relies upon the fact that mutations acquired by an individual can be early acting or late-onset. Early acting mutations manifest themselves during the period where individuals are reproductively active, so that whatever effect the mutation has will be acted upon by natural selection. However, natural selection is ‘blind’ to late-onset mutations. Mutations that reveal themselves later in life cannot be retroactively selected against, if reproduction has already occurred, and passed on to the next generation. Therefore, the force of selection diminishes with age and reproductive decline. This phenomenon is known as the selection shadow. (Beyond A Shadow Of A Doubt!)
The selection shadow is the diminished force of selection with age and reproductive decline. Source: Fabian and Flatt, 2011
This theory contributes to why it is so difficult to breed animals in captivity till ages later than they’d naturally experience in the wild. When their environment changes and survival risks are lifted, such as in captivity, animals can live to an age where accumulated late-onset mutations can manifest themselves. This effect is exaggerated by mutation concentration due to inbreeding in these small populations. Conservationists have great difficulty dealing with the elevated level of these age-related diseases, which are not seen at such rates in the wild. A similar phenomenon has been observed with the rise of neurodegenerative diseases in human populations, co-occurring with our drastic increase in lifespan over the past century.
The mutation-accumulation model was refined in 1957 by the American evolutionary biologist George C Williams, when he elaborated further on the relationship between early acting and late-onset mutations. A mutation can have multiple effects that present themselves differently in different tissue types, or different stages of life, a genetic phenomenon known as pleiotropy. When the mutation has opposing effects on fitness at different life stages, for example during early and late life, it is known as antagonistic pleiotropy. Williams postulated that if a mutation has a beneficial effect on survival and reproduction in early life, but a negative aging-related impact in later life, selection will act upon the early benefit and enrich this mutation in the population. This antagonistic-pleiotropy model presents aging as a maladaptive byproduct of selection for survival and reproduction during youth.
The question of why we die of old age is a complex one, and the details are beyond the scope of this book. In addition to particular reasons, some more general ones have been proposed. For example, one theory is that senility represents an accumulation of deleterious copying errors and other kinds of gene damage which occur during the individual's lifetime. Another theory, due to Sir Peter Medawar, is a good example of evolutionary thinking in terms of gene selection. Medawar first dismisses traditional arguments such as: ' Old individuals die as an act of altruism to the rest of the species, because if they stayed around when they were to decrepit to reproduce, they would clutter up the world to no good purpose.' As Medawar points out, this is a circular argument, assuming what it sets out to prove, namely that old animals are too decrepit to reproduce. It is also a naive group-selection or species-selection kind of explanation, although that part of it could be rephrased more respectably. Medawar's own theory has a beautiful logic. We can build up to it as follows.
We have already asked what are the most general attributes of a 'good' gene, and we decided that 'selfishness' was one of them. But another general quality that successful genes will have is a tendency to postpone the death of their survival machines at least until after reproduction. No doubt some of your cousins and great-uncles died in childhood, but not a single one of your ancestors did. Ancestors just don't die young!
A gene that makes it possessors die is called a lethal gene. A semi-lethal gene has some debilitating effect, such that it makes death from other causes more probable. Any gene exerts its maximum effect on bodies at some particular stage of life, and lethals and semi-lethals are not exceptions. Most genes exert their influence during foetal life, others during childhood, others during young adulthood, others in middle age, and yet others in old age. (Reflect that a caterpillar and the butterfly it turns into have exactly the same set of genes.) Obviously lethal genes will tend to be removed from the gene pool.But equally obviously a late-acting lethal will be more stable in the gene pool than an early-acting lethal. A gene that is lethal in an older body may still be successful in the gene pool, provided its lethal effect does not show itself until after the body has had time to do at least some reproducing.For instance, a gene that made old bodies develop cancer could be passed on to numerous offspring because the individuals would reproduce before they got cancer. On the other hand, a gene that made young adult bodies develop cancer would not be passed on to very many offspring, and a gene that made young children develop fatal cancer would not be passed on to any offspring at all. According to this theory then, senile decay is simply a by-product of accumulation in the gene pool of late-acting lethal and semi-lethal genes, which have been allowed to slip through the net of natural selection simply because they are late-acting.
The aspect that Medawar himself emphasizes is that selection will favour genes that have the effect of postponing the operation of other, lethal genes, and it will also favour genes that have the effect of hastening the effect of good genes. It may be that a great deal of evolution consists of genetically-controlled changes in the time of onset of gene activity.
It is important to notice that this theory does not need to make any prior assumptions about reproduction occurring only at certain ages. Taking as a starting assumption that all individuals were equally likely to have a child at any age, the Medawar theory would quickly predict the accumulation in the gene pool of late-acting deleterious genes, and the tendency to reproduce less in old age would follow as a secondary consequence.
https://io9.gizmodo.com/what-is-the-evolutionary-advantage-of-death-743044300 We are all getting older. The average human lifespan has doubled in the past one hundred years, prompting claims that 60 is the new 40 and 50 the new 20. But decade relabelling aside, we are still ageing in much the same way we always did: as we grow older, our risk of dying increases dramatically. Once the average British male hits their late 20s, their (initially small) chances of passing away will double with every decade that follows.
As an aside, one of the good features of this theory is that it leads us to some rather interesting speculations. For instance it follows from it that if we wanted to increase the human life span, there are two general ways in which we could do it. Firstly, we could ban reproduction before a certain age, say forty. After some centuries of this the minimum age limit would be raised to fifty, and so on. It is conceivable that human longevity could be pushed up to several centuries by this means. I cannot imagine that anyone would seriously want to institute such a policy.
Secondly we could try to 'fool' genes into thinking that the body they are sitting in is younger than it really is. In practice this would mean identifying changes in the internal chemical environment of a body that take place during ageing. Any of these could be the 'cues' that 'turn on' late-acting lethal genes. By simulating the superficial chemical properties of a young body it might be possible to prevent the turning on of late-acting deleterious genes. The interesting point is that chemical signals of old age need not in any normal sense be deleterious themselves. For instance, suppose that it incidentally happens to be a fact that a substance S is more concentrated in the bodies of old individuals than of young individuals. S in itself might be quite harmless, perhaps some substance in the food which accumulates in the body over time. But automatically, any gene that just happened to exert a deleterious effect in the presence of S, but which otherwise had a good effect, would be positively selected in the gene pool, and would in effect be a gene 'for' dying of old age. The cure would simply be to remove S from the body.
What is revolutionary about this idea is that S itself is only a 'label' for old age. Any doctor who noticed that high concentrations of Stended to lead to death, would probably think of S as a kind of poison, and would rack his brains to find a direct causal link between S and bodily malfunctioning. But in the case of our hypothetical example, he might be wasting his time!
There might also be a substance Y, a 'label' for youth in the sense that it was more concentrated in young bodies than in old ones. Once again, genes might be selected that would have good effects in the presence of Y, but which would be deleterious in its absence. Without having any way of knowing what S or Y are - there could be many such substances - we can simply make the general prediction that the more you can simulate or mimic the properties of a young body in an old one, however superficial these properties may seem, the longer should that old body live.
I must emphasize that these are just speculations based on the Medawar theory. Although there is a sense in which the Medawar theory logically must have some truth in it, this does not mean necessarily that it is the right explanation for any given practical example of senile decay. What matters for present purposes is that the gene-selection view of evolution has no difficulty in accounting for the tendency of individuals to die when they get old. The assumption of individual mortality, which lay at the heart of our argument in this chapter, is justifiable within the framework of the theory. (The Selfish Gene)
"We're shaped for reproduction, not health. When it comes to sex and reproduction, we do things that are bad for our health and longevity, but good for our genes"
https://www.amazon.com/Mismatch-Lifestyle-Diseases-Peter-Gluckman/dp/0199228388 I'M NOT DONE WITH THIS BLOG POST. THE PASSAGE I TAKE FROM THIS BOOK CONCERNING THE EVOLUTION OF DEATH WILL BLOW YOUR MIND. JUST WAIT UNTIL I WRITE IT OUT (WHICH I SHOULD DO BY TOMORROW). WE'RE GENETICALLY EVOLVED TO REPRODUCE, NOT LIVE LONG! One cost of living longer has been the rapid rise in the occurrence of diseases of degeneration and of middle and old age. These include cancer, diabetes, neurodegeneration, heart disease, and conditions which some would regard as a normal part of ageing, such as osteoporosis, osteoarthritis, and a decline in mental ability. For some of these conditions, their appearance as we age is due to the failure of repair and maintenance systems. However there is ongoing debate amongst gerontologists about the specific processes involved in ageing. Either the body may simply not be able to afford additional maintenance; or the impact of progressive environmental insults on cellular function accumulates; or there is some inherent process of senescence that gives a finite life to particular tissues because they have not been designed to last. While there are multiple theories of the biology of ageing they all basically come down to this cluster of possibilities. We favor the group of theories (there are several variants on the theme) that there is a trade-off between the lifetime investment in growth, reproductive, and repair systems. According to this theory those species and individuals which anticipate a short life invest less in repair and more in early reproduction and vice versa.
Once an individual has ceased reproduction and support of their progeny, there can be no selection pressures acting on them (with one possible exception we will shortly consider) and therefore there will not be selection against the inherent process of ageing. Females have an absolute end to reproduction at menopause but reproductive capacity declines from about the age of 35, probably because the eggs which were all formed in a woman's fetal life are growing old and are less viable. In males reproduction is possible throughout life after puberty but, while we can only speculate about Paleolithic social structures, it is probable that male reproductive opportunity also declined with age. There are some important clues to support this idea. For example males are bigger than females and this can be taken as evidence of competitive selection for size in males. This implies that the larger and stronger males, and therefore probably younger, healthier males, had mating dominance. So again selection would not have worked to the advantage of the older male. The older male past his mating prime may have been like the old male lion, banished from the pride by the younger male, doomed to a lonely death. Alternatively, as in some primate species such as the gelada baboon, the older male remains with the colony but makes no attempt to re-enter the mating game. In some pre-colonial societies the old and infirm were intentionally allowed to die. Yet in other societies such as the Nicobarese and some indigenous American and Australian peoples, the aged were treated with the greatest of respect. In some societies old age mattered greatly in that it may have conferred the capacity of the clan to pass information through cultural inheritance with greater fidelity. Wisdom, experience, and knowledge could be important to survival under extreme condition such as drought and this was most likely to be held in the memories of the oldest. Recent studies in the African elephant, which lives in small herds dominated by a matriarch, show that the presence of the grandmother elephant confers collective memory on the herd. It is used for example to recognize other herds of elephants as non-threatening even when previous contact with them has been infrequent.
We evolved with a life-history strategy that was characterized by living in small clans at the forest edge, caring for big-brained, slowly maturing offspring. This meant that we had to space our pregnancies out appropriately. But even once her children were through infancy the Paleolithic mother had still to support her older children until they were fully independent. A high percentage of offspring survived to reproduce - about 50 percent, one of the highest in the animal kingdom. This strategy of having a small number of children and high parental investment led to social structures where there was some stable bonding between mother and father so that the father was also involved in child support. This was not simply altruistic because it also ensured that his genetic endowment was protected. Although the environment we lived in generally led to death from trauma, childbirth, or infection by the end of the fourth decade, in any event by then our reproductive role was effectively over. So it is reasonable to assume that we evolved living in an environment where reproduction was largely complete by age 35 years of age and life expectancy was not much longer. In such a system there would be little evolutionary pressure, and in fact potentially significant cost, to having repair systems that were effective for a period of many subsequent decades, although there would have always been some individuals who survived longer. This may explain why our repair and maintenance systems become less effective in middle and old age, and it is this decline that creates one of the mismatches of ageing. We now enjoy an average a lifespan more than twice that anticipated by our Paleolithic ancestors. The repair systems are not designed for this, and they cannot cope.
I'LL FINISH THE REST OF THIS TOMORROW. PLENTY MORE TO COME.
"Contrary to the widespread assumption that hunter-gatherers die young, foragers who survive the precarious first few years of infancy are most likely to live to be 68 to 78 years old...not far from the life expectancy in the US which is between 76 and 81" https://harvardmagazine.com/2020/09/features-active-grandparenting

 CARTAUCSD
CARTAUCSD